
森林と樹木の健康:2 樹液流の持続性
神戸大学 森林資源学研究室 黒田慶子 Kuroda, Keiko
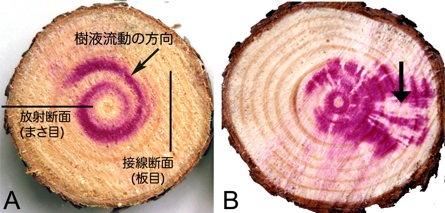
図-3 酸性フクシン注入によるクロマツ木部の通導経
路の検出
A:健全時の螺旋状の上昇.
B:線虫接種3週間後にみられた水分通導の乱れ.矢
印:通導が停止した部位
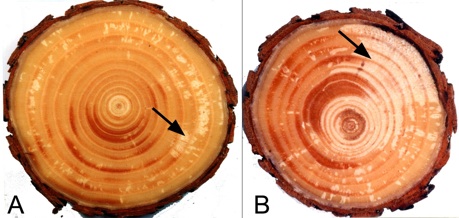
図-4 マツ材線虫病罹病アカマツにみられる仮道管の
排水(エンボリズム)
A:接種3週間後に観察された白い斑点(矢印).
B:接種6週間後、萎凋開始直前の通導停止状態
矢印:排水して気体に置き換わった部位
針葉樹の仮道管の直径は20-30µm程度である(図−1A, B)(佐伯 1982,島地,伊東
1982).水不足や外傷によって通導組織で局所的に気泡が発生した場合,仮道管の壁孔にある膜は細かい網目(直径1µm以下)になっているので(図
-2),気泡は隣の仮道管に広がりにくく,排水は狭い範囲に押さえられる.一方,広葉樹にある道管には大きな穴
(せん孔)があり,気泡が発生した場合に広
範囲に広がって水の流れが止まりやすい.道管は水を運ぶ効率はよいが,通導機能が停止しやすいという欠点がある.ニレやハリエンジュ,コナラなどの材は環
孔材と呼ばれ,直径が0.3㎜(300µm)もある大きな道管が同心円状に並んでおり,その他に小径の道管が多
数分布する(図-1C).大径道管は形成さ
れた年の数ヶ月から数年程度しか機能しないが,小径の道管心材になるまで何年も機能して水をあげ続ける.
筒状の通導要素と直交する放射組織の柔細胞は(図-1B),数年から時には十数年以上生き続けることができる.この細胞は物質の代謝や糖(でんぷん粒)の
貯蔵,防御反応,老廃物の貯蔵を担う(島地ら1976).これまで長らく,木部樹液の流動は単に通導要素の筒の
中を水が通る「物理現象」であると考えられ
てきたが,近年,放射組織の柔細胞が仮道管や道管内の水の動きに重要な役目を果たすという説が有力になってきている.


樹木医学研究 11:83-88,2007
樹木医学の基礎講座
2. 木部樹液の動きと樹木の健康
I.はじめに
生きている樹木の木部組織(材の部分)を揚がる水は
「木部樹液」と呼ばれ,人間の血液に匹敵するほど重要である.
人間と水については医学的研究が進み,脱水症状など日常的に話題になることも多い.しかし樹木の水分吸収や利用の仕組みについては,樹木を扱う仕事をして
いても,今ひとつわかりにくいという印象がある.もちろん,水がなければ枯れることはわかっているが,どのよう
な場面で水不足がおこるのか,水不足の場合
に樹木はどのような生理状態になるのかなど,わからないことが多い.たとえば,樹木を人為的に枯らす手法として知られる「巻枯らし」では,なぜ枯れるので
あろうか.樹皮を環状に剥いだ樹木が枯れるのは,実は「辺材の乾燥が進んで水が揚がらなくなる」ためであるが,
意外にも「形成層が死ぬから」「養分が届か
なくなるから」「通導組織が傷つくから」といった誤った認識がある.大学の林学関連分野でさえ樹木の組織構造と機能については十分な講義が行われないの
で,当然起こる誤解であろう.
樹木病害関連の研究報告では,樹木に水分供給が足りない状態をさして「水ストレスがかかる」と表現することがよくある.私たちは,そのような条件下で病気
の感染や発病が起こりやすいことを経験的に知っている.また,「萎凋病」をおこす病原体に感染すると,木部樹液
の流動が悪くなり,枯死するというしくみに
なっている.その他にも,根腐れのために水分が吸収できなくなって枯れる場合もある.このような「水分」の関係で樹木に起こるトラブルについては一部を除
いて充分に研究が進んでいないが,いくつかの事倒について解説したい.

図-2 健全な樹木の仮道管にみられる蒸散によるエン ボリズム(排水)
図-1 木部の組織構造
A: クロマツ横断面、B:クロマツ放射断面、C:コナラ横断面
IV.気象変動によるトドマツの集団枯死
北海道の道東地方(釧路,阿寒など)では,人工林
のトドマツが集団で枯れることがあり,1989年の初夏に70年生林分が200ha枯死した.萎凋にかかわる昆
虫の侵入や微生物の感染はなかった.被害の発生がこれだけ広範囲で一斉である場合には,生物害とは考えにくく,
気象の変動との関係が注目される.枝先の凍結壊死はないので,低温による枯死(凍死)でないことは明らかであっ
た.葉枯れが見られる個体の主幹では木部含水率が低かった.色素液を樹幹下部から注入したところ,通導機能が低
下しており色素液が少ししか上がらなかった.また,葉枯れを起こしている枝の根元を色素に浸すと,木部の一部し
か染色されず,断面の大半は通導が停止していること,通導停止部位周辺では皮層と形成層の壊死が起こっているこ
とが確認された(図-6).その時期に土壌水分は充分にあったので,何らかの原因で通導停止が起こったものと推
測した(黒田2000).
道東地方では冬季の気温が非常に低く,真冬日(一
日の最高気温が0℃を超えない)が続く.師部や形成層の生きている細胞は冬季には耐凍性があって内容物は凍らな
いと言われているが,純水に近い木部樹液は-6℃程度で凍ることが確認されている.2月頃は木部樹液が完全に凍
結しており,3月下旬から4月に気温が上昇して日中の気温が0℃以上になると,樹液は融解し始める.主幹下部や
根の樹液が凍結している時期に日の当たる葉で蒸散が起こった場合,樹液上昇はなく,梢端や主幹上部から下方に向
かって乾燥が進む.2月に暖かい日が続いた1999年には,多数の仮道管が空になったまま1ヶ月以上過ぎ,4月
以降に樹液や土壌がすっかり説けて根が吸水できる状態になった時には,通導機能が回復できなくなっていたと推測
された.樹液の通るルートが極端に減ったため,蒸散が活発になる初夏に樹冠への水分供給が決定的に不足し,急激
な葉枯れや枯死へと進行した(図−7).この大面積の枯損は1999年春の異常気象のせいであろうと考えられる
が,小面積や数本単位の枯損は以前から知られている.道東は本来トドマツの天然分布地域ではなく,昔からトドマ
ツの植栽は望ましくないと言われていたらしい.適地適木の教えを軽んじた結果起こった現象である.

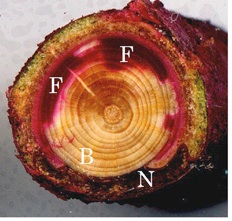
図-5 ナラ類集団枯死木の横断面
A:健全木の心材と辺材,
B:被害木辺材の変色(傷害心材),
C:変色部の放射断面(顕微鏡写真).
伸長した菌糸が柔細胞に侵入している.
V.おわりに
萎凋を起こす病気や気象変動について数例しか紹介
できなかったが,他にも萎凋現象はいろいろある.暗色枝枯病に繰り返し感染したスギやヒノキでは,通導できる木
部が極端に減少し,夏の乾燥(例年にない干ばつ)が重なると,梢端枯れや枯死が増える.ま
た,Ceratocystis属菌の感染によるエゾマツの青変病の場合,数か所以下の病原菌接種では通導の停止
が部分的にしか起こらず,人為的に枯らすのは難しい.しかし自然感染ではキクイムシが多数侵入して病原菌を「多
点接種」することになり,通導停止範囲が広がって枯れる.針葉樹と広葉樹では,水分通導要素のサイズや配置が異
なるので,萎凋の起こり方や枯れ方には違いがある.また,樹木の種のレベルで水不足に対する耐性が異なり,これ
が病気に対する「感受性の差」として現れることがある.
樹木に穴をあけて枯らそうとしても,簡単には枯れ
ないものである.破壊された通導要素が多くなければ,その部位を迂回して木部樹液は上昇できる.樹木が枯れるか
生き抜くかは,病原体の活動,樹木の生理,および環境条件とのバランスで決まり,どちらに転ぶか微妙なである.
樹木医学は,患者である樹体の組織構造や機能,感染後の異常生理に関する知識をもとにして診断するシステムがで
きておらず,「顔色を見て診断する」民間療法の段階に留まっていると言わざるを得ない.人間の病気の診断とくら
べるとまだまだこれからである.枯れた樹木から糸状菌が検出された場合,その菌がトラブルの真の原因であるの
か,枯れかけてから侵入して子実体を作ったのか,その見極めができていないことがよくある.病原体が明らかな場
合でも,なぜその病原体が活動する状態になったのか,樹木の生理や環境因子に目を向けて,きちんと情報を得て診
断する必要がある.罹病や発病の促進要因について研究が進めば,診断に留まらず,将来の枯死を減らすような予防
医学的な対策が可能になるであろう.
水分
通導や萎凋病に関する解説はhttp://www2.kobe-u.ac.jp/~kurodak/に掲載しており,著者の論文(PDFファイル)はダウンロード可能であ
る.
引用文献
1.黒
田慶子(1999)樹木医学,2.2樹木の構造と機能(鈴木和夫編),朝倉書店 57-82.
2.黒
田慶子(2000)道東トドマツ壮齢人工林に発生した集団枯損 ---水分通導阻害がなぜ起こったのか
---,森林保護 276:12-14.
3.Kuroda,
K. (2001) Responses of Quercus
sapwood to infection with the pathogenic fungus
of a new wilt disease vectored by the ambrosia
beetle Platypus
quercivorus. J. Wood Science
47:425-429.
4.黒
田慶子(2003)マツ樹幹内で起きていること ---マツ材線虫病の発病機構と抵抗性に関する研究よ
り--- 森林防疫 52:19-26.
5.黒
田慶子(2004)森林保護学:3.1.1
樹木の構造と機能を測る(鈴木和夫編著),朝倉書店 86-94.
6.Kuroda,
K., Kuroda, H. and Ann M. Lewis (2000) Detection
of embolism and acoustic emissions in tracheids
under a microscope: incidence of diseased trees
infected with pine wilt, in "New Horizons in
Wood Anatomy", Y.S. Kim ed., New Horizons in
Wood Anatomy, Chonnam National Univ. Press
Kwangju, Korea, 372-377.
7.黒
田慶子,山田利博(1996)
ナラ類の集団枯損にみられる辺材の変色と通水機能の低下.日本林学会誌 78(1):84-88.
8.佐
伯 浩(1982)走査電子顕微鏡図説 木材の構造,218pp,日本林業技術協会.
9.島
地 謙,伊東隆夫(1982)図説木材組織,176pp,地球社.
10.島地 謙,原田
浩,須藤彰司(1976)木材の組織,291pp,森北出版.
11.Zimmermann,M.H. (1983) Xylem
structure and the ascent of sap. 143 pp.,
Berlin: Springer-Verlag.
図-6 トドマツの部分枯損個体の枝に見られる通導阻害
F:酸性フクシンにより染色された通導部,
B:通水阻害部位,N:形成層の壊死
図-7 凍結時の蒸散による通導阻害の発生機構:仮説

II.健康な樹木の水分通導
最初に,健康な樹木の水分通導について述べる.樹
木内の水分移動について理解するには樹木組織学の知識が必須であるが,ここでは詳しく解説できないので,教科書
等(佐伯1982, 島地ら1976, 黒田 1999, 2004)を参照していただきたい.
土壌中の水はミネラル類とともに根毛から吸収され,木部に運ばれる.高分子や水に不溶の物質は根から吸収されない.根にも木部と師部があり,幹と同様に肥
大成長するが,髄がないのが特徴である.水分の通路となる通導組織は中空の筒(道管,仮道管)の集合で(図
−1),その形状や分布,葉までの水の揚がり方
は樹木の種類により特徴がある.健康な樹木では,通導組織はほとんど混入物のない水で満たされている.根の木部の水は,葉から水が蒸発する(蒸散)時に発
生する「テンション(張力)」と水分子の「凝集力」によって数十メートル以上も上昇し,梢の先まで達することが
できる.この概念は「凝集力 説」"cohesion-tension
theory"として1980年代に確立された(Zimmermann
1983).通導システムに不具合が生じたとき,樹木は萎れたり枯れることになる.
蒸散が活発なときに土壌中の水分が不足していると,木部内では樹液にかかる張力が非常に高くなり,水分子間のつながりがその力に耐えられなくなる.すると
樹液中に気泡が突然発生し,道管や仮道管などの通導要素は気体で満たされる(図-2).わかりやすくたとえる
と,「減圧下で水が沸騰する」イメージであ
る.この現象はエンボリズムあるいはキャビテーションと呼ばれ,気泡と同時にアコースティックエミッション(AE,超音波の一種)が発生する.樹幹組織内
での気泡発生は木部切片を用いて顕微鏡下で観察する方法があり,AEと気泡発生を同時に記録した映像をhttp://www2.kobe-u.ac.jp/~kurodak/embolism_j.htmlに
掲載した(Kuroda et al. 2000).
強い蒸散によって樹液流が途切れる現象は,天気が良ければ毎日どの樹木でも起こっている.しかし夜間あるいは降雨により樹液にかかる張力が弱まると,空に
なった道管や仮道管にはまた水が流入する.水分の上昇が回復するので,樹木は簡単に枯れることはない
(Zimmermann
1983).しかし,夏に降雨のない日が続くと通導組織の中の水は減る一方となる.木部がカラカラになった状態がある期間(樹種により異なる)続くと,そ
のあとで水を与えても樹液の流動は回復しないことがわかっている.乾燥害といわれる枯れはこのように起こってい
ると考えられる.近年,北海道でトドマツ林
に発生した集団枯死は,土壌や樹幹が凍っている早春に蒸散が起こり,木部が乾燥したことが原因であろうと推定している(黒田2000).詳細は後述する.
針葉樹の場合,根からの吸水は蒸散による引っ張りの力にのみ依存しているが,広葉樹(双子葉植物)ではヘチマ水の採取に見られるように,根が積極的に水を
取り入れる力を持つといわれている.しかし「根圧」がどこでどのように発生するのか,よくわかっていない.この
現象とは別に,広葉樹では葉が展開していな
い早春に木部樹液が上昇する現象が見られる.気温が0℃前後で変動し樹液が凍結融解を繰り返す時期にシラカバやカエデの樹液(メープルシロップの原料)が
採取できる.冬季に糖濃度が高まっていた細胞が早春に吸水するため,樹液流動が起こると説明されているが,科学
的な証明はなされていない.
III.萎凋病に感染した樹木の通導停止
微生物に感染して「突然枯れる」病気を一般に萎凋
病と呼んでいる.世界的に有名な病気としては,マツの材線虫病,針葉樹類(トウヒ,マツなど)の青変病,ニレ立
枯病,ナラ萎凋病(oak
wilt)があり,近年日本で被害が拡大している「ナラ類の集団枯死」もその一つである.突然枯れる原因として,「水(木部樹液)が揚がらなくなる」こと
がこれらの病気に共通している.欧米ではニレ立枯病について研究が進められ,セラトウルミンという毒素の関与が
報告されている.日本ではマツ材線虫病が猛威をふるっているため,枯死のメカニズム解明を目指した研究が精力的
に進められ,多くの成果が出ている.ナラの集団枯死については,菌の感染と通導阻害に関する知見が得られつつあ
る.このような萎凋病感染で起こる水分通導停止について解説する.
1.マツ材線虫病
病原体であるマツノザイセンチュウ(以下線虫)
は,成虫の体長が1㎜ほどで,マツの皮層や木部にある樹脂道の中を移動し,樹脂道を取り囲むエピセリウム細胞や
柔細胞の内容物を酵素分解して口針で摂取する.日本のアカマツ・クロマツは線虫を接種すると7〜9割が枯死す
る.しかしこの線虫がマツの細胞を直接食い荒らして枯らしているのではいない.また,毒素がマツを枯らすという
説があったが,毒で細胞が多数死ぬような現象は観察されていない.
樹脂道は水分通導とは無関係で,外敵の侵入を防御
するための樹脂(松ヤニ)の分泌に関わる組織である.線虫が樹脂道をつたって樹幹内を動き回る速度は非常に速
く,最大で150㎝/日である.木部樹液の上がる様子は,樹幹の下部に穴をあけて色素液(酸性フクシン水溶液な
ど)を注入してから伐到すると肉眼で認識することができる.健康なマツの樹幹では樹液は螺旋状に上昇しているが
(図−3A),線虫を接種したアカマツやクロマツでは,2〜3週間後には水分通導できない仮道管が急激に増加
し,樹液はその部位を迂回して上昇するため,赤い色素は螺旋状のパターンを示さなくなる(図−3B)(黒田
2003).
線虫が樹脂道内を移動して細胞を刺激すると,マツの
細胞は二次代謝物質(テルペン,スチルベン,フェノール類など)を生産し,アカマツやクロマツではαピネンなど
揮発性テルペン類が急激に増加する(黒田2003).他方,樹脂の主成分である樹脂酸など分子量の多い物質の生
産は減少し,樹脂の分泌は停止する.材線虫病の検査に樹皮を剥いで樹脂の出方を見るのは,樹脂生産の停止を
チェックしているのである.線虫接種の2週間後ころから,樹幹横断面では白い斑点が出現する(図-4A).これ
が「排水して気体に置き換わった部分」で,この部位を樹液は通れない.斑点はその後拡大し,4〜6週間後ころに
は乾燥がすすみ(図−4B),葉の萎れや変色など外見的病徴も明瞭になる.雨が降っても,大量に灌水しても,白
く乾いた部分が縮小することはない.これが健全な樹木のエンボリズムと異なる点である.テルペン類など疎水性の
物質が細胞壁や壁孔に付着しているため,水が供給されても空の仮道管内には流入しにくいと考えられる.
このように,材線虫病感染木では「回復不能の排水」が起こっている.線虫は移動速度が速いため,感染木では時
には1週間程度で樹幹全体に線虫が分布し,その各所で通導停止が起こる.渇水期の夏季,しかも蒸散が活発な時期
に通導可能な部位が急激に減少するため,枯死に至るまでの時間が非常に短くなる.マツは外敵に対する防御物質
(二次代謝物質)の生産を盛んに行い,同時に自ら樹液の流動を止めてしまうという皮肉な結果となっている.線虫
は生産された二次代謝物質の影響をそれほど受けないようである.通導の停止に関わる物質はテルペン類以外にもあ
るかも知れないが,今のところ具体的な物質名はあがっていない.マツノマダラカミキリはテルペンなど増加した揮
発性物質に誘引され,枯死が近いマツを見つけて効率よく産卵する.
2.ナラ類の集団枯死
1980年代の後半からミズナラやコナラ,時には
シイやカシ類が夏季に集団で枯れる現象が目につくようになった.病原体は糸状菌Raffaelea
quercivoraであり,養菌性の昆虫(アンブロシアビートル)カシノナガキクイムシがこの菌を元気なナラ類に感染させる.この甲虫は萎凋しかけたナ
ラ類の辺材に長いトンネル(孔道)を掘って子育てを行い,翌年春に菌を体につけた成虫が脱出する.
樹幹内に持ち込まれた菌は生きている放射組織柔細
胞に侵入することができる(図5C) (Kuroda
2001).菌糸の周辺の柔細胞ではマツの場合と同様に防御反応として二次代謝物質の生産が活発になり,それが道管内に放出される.無染色
の切片を顕微鏡
で観察すると,油性の物質が道管内に付着しているのが観察される.二次代謝産物が多量に生産された部位では木部
組織が褐色〜黒褐色に変色する(図5A,
B).この変色は傷害心材や病理的心材とも呼ばれ,通常の心材と同様に通導機能は全く失われている
(黒田,山田
1996).通導が停止した大径道管にはチロースという風船状の物体の充填が認められるが,チロースが通導を止めたのではなく,通導が止まった後に形成さ
れたものである.
菌糸は孔道を利用して迅速に分布拡大できるので防
御反応の効果は小さく,辺材全体が短期間で変色し,通導停止することになる.ただし,マツ材線虫病の揚合のよう
な白い斑点状の排水部位は観察されない.広葉樹の道管は分布密度が針葉樹の仮道管より低く,排水して気体に置き
換わった状態になったかどうか,肉眼では認識できない.ミズナラやコナラの大木が感染後1〜2ヶ月で枯死する.
健全な樹木で心材化の際に道管や仮道管が通導しなく なる仕組みについては,これまで充分には説明されていなかった.しかしR. quercivoraに感染したナラ類で辺材が変色し傷害心材の形成が進む過程では,二次代謝産物が柔細胞から放出されて通導機能が停止し,さらに柔細胞 類の壊死という現象がほぼ同時に起こっていることが明らかになった (Kuroda 2001)