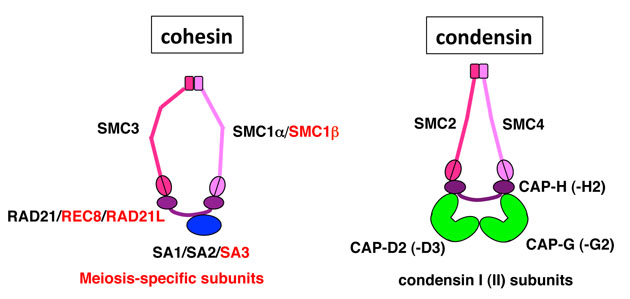
我々ヒトを含む哺乳類の体を構成する細胞は、染色体のセット(ゲノム) を2組み持っています。この2組みのゲノムは、受精の際に精子と卵子から 1組みずつ持ちもまれたものです。受精卵は、細胞分裂のたびに染色体を正 確に複製し分配するので、1つの個体を構成する体細胞は、一部の例外を除 いて、全く同じ2組みのゲノムを持っています。精子や卵子のもととなる精 母細胞や卵母細胞は、減数分裂により染色体の数を半減します。そうするこ とによって、世代交代の際の、染色体数の倍加を防ぎます。また、この過程 においてもう一つ重要なことは、形成される配偶子が遺伝的な多様性を持つ ことです。これは、交差を伴う相同染色体の組換えによりハイブリッド型の 染色体が生み出されることと、父母由来の相同染色体が分配されるときに幾 通りもの染色体の組合わせができることによります。 それでは、減数分裂は体細胞分裂とどこが違っているのでしょうか?減数 分裂と体細胞分裂では、以下の2点に大きな違いがあります。 【1.細胞周期】体細胞分裂においては、DNA複製期(S期)と分裂期(M期) が交互に繰り返され、細胞は1サイクルごとに倍増します。一方、減数分裂 においては、S期の後にM期が2回連続して起こります。また、雌の減数分裂 は他にも特徴があり、卵母細胞は、第一減数分裂前期で長い期間(ヒトの場 合では数十年に及ぶ)休止します。さらに、減数分裂を再開後も、第二減数 分裂中期で再び分裂を休止し、そこで排卵され、精子と受精します。 【2.染色体の動態】体細胞分裂における染色体の分配は、S期に複製された 染色体(姉妹染色分体)が分裂後期に分離します。減数分裂では、まず第一 減数分裂前期に相同な染色体がペアをつくり、組換えを行います。その結果、 相同染色体間に結合が生じ、二価染色体が形成されます。第一減数分裂の後 期には、相同染色体が分離しますが、このときには、姉妹染色分体はセント ロメア部分で接着が維持されます。続く、第二減数分裂の後期において、セ ントロメアの接着が解消されて、姉妹染色分体が分離します。 体細胞分裂においても減数分裂においても、染色体の接着には、コヒーシ ンと呼ばれるタンパク質複合体が中心的な役割を果たします。しかし、減数 分裂の時期には、特異的なサブユニットがいくつか発現し、それが減数分裂 に特徴的な染色体動態の制御に重要であることがわかってきています。一方、 コンデンシンと呼ばれるタンパク質複合体は、染色体の凝縮・分離に寄与す ることが知られています。哺乳類ではコンデンシンIとコンデンシンIIと呼 ばれる2つのタイプが存在し、減数分裂においてもその両者が発現している が、減数分裂における個々の役割はまだ明らかとはなっていません。また、 興味深いことに、コヒーシンもコンデンシンも、染色体の接着や凝縮という よく知られた役割以外に、遺伝子発現の制御やヒトの遺伝疾患との関連が最 近明らかとなってきています。 我々は、多彩な染色体機能に関わるコヒーシンやコンデンシンに着目して、 減数分裂を制御する分子メカニズムを明らかにすることを当面の課題として います。哺乳類の生殖細胞を実験材料とした、このような基礎研究は、家畜 の繁殖やヒトの生殖医療などの分野において新規な基盤技術を開発する上で、 非常に重要な知見を与えてくれるものと期待しています。
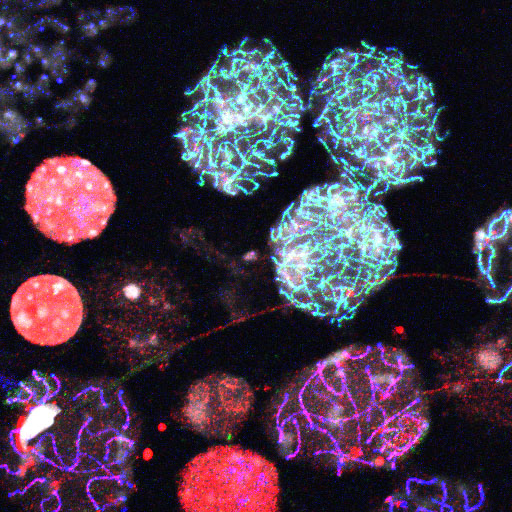
マウス精巣細胞の免疫蛍光写真;シナプトネマ複合体SYCP3(青)、
コヒーシンRAD21L(緑)、コヒーシンRAD21(赤)、DNA(銀)
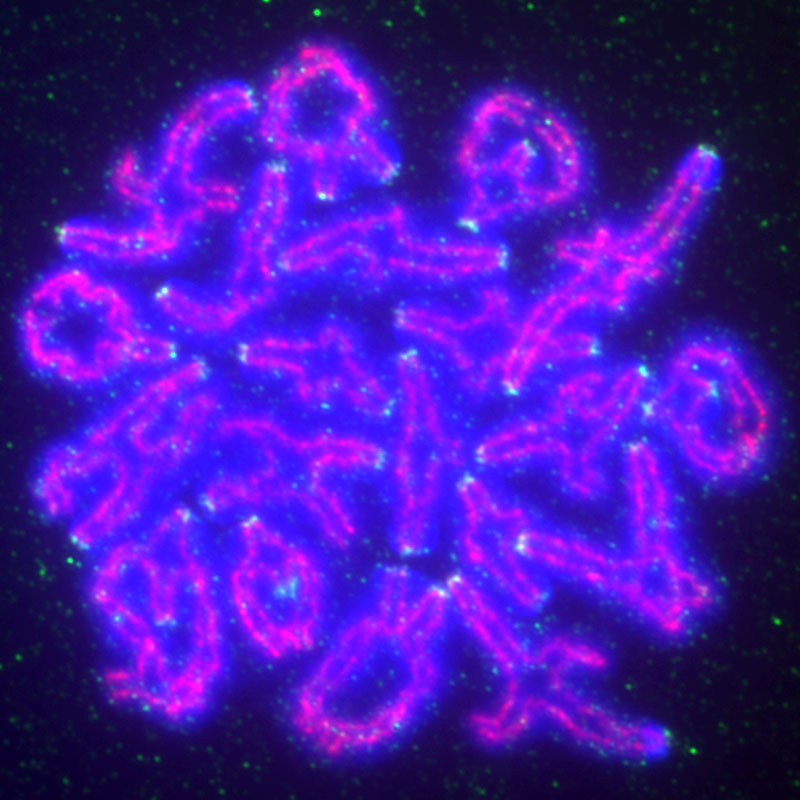
マウス卵母細胞の二価染色体の免疫蛍光写真;DNA(青)、
セントロメア(緑)、コンデンシンII(赤)