神戸大学農学部 森林資源学研究室 黒田慶子
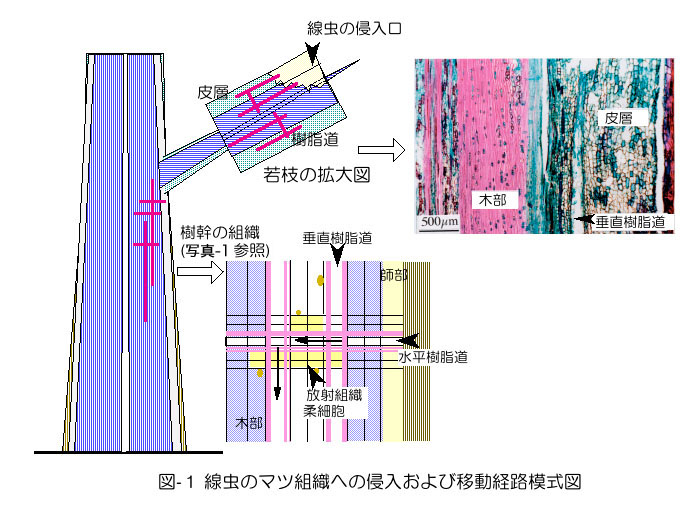
マツ材線虫病の発病と枯死のメカニズムは,かなり詳しく説明できるようになりましたが、「よくわ かっていない」と思われることが多く,「マツノザイセンチュウ(以下線虫)は病原体でないかも知れない」と疑いを持たれる原因ともなっています.その誤解 をとくために、ここでは「線虫の感染からマツが萎凋するまでの道筋」について基本的な事柄を整理して解説します.幹の外から見えない現象がイメージできる ように,マツの組織の立体構造や機能について,図や写真で説明しています(図-1,写真-1).高校で習った生物の知識でわかっていただけると思います が、草と木の組織とその機能はかなり違っています。ここを読む前に、「樹 幹組織の成り立ち」の解説をご覧ください。
線虫は、若い枝の傷(マツノマダラカミキリの噛み傷)からマツの組織に侵入し(図-1)、樹脂道の中を通って樹幹の本体の方に移動します。線虫の長さは 1mm程度で、移動しながらマツの生きている細胞の内容物を吸い取り、養分としま す。マツの2〜3年生程度までの若枝は、木部(材)の外側には皮層と表皮があり、内樹皮(二次師部)は少しかありません(黒田 1999)。枝が太くなるにつれて皮層は剥がれ落ち、外樹皮・内樹皮と木部という構造になります。枝の皮層の中の線虫は水平樹脂道を通って木部に移動し、 それから幹の中を移動することになります。この点について理解 を深めるには、樹木の組織学の知識が必要になります。
2.目に見える変化の前に樹幹内で起こる「水分 通導のトラブル」
線虫が感染したクロマツやアカマツに最初に見られる外観の変化は旧葉(1年以上前からある葉)の変色です。しかし樹幹の中ではそれ以前に大きな変化が起
こっています。感染から病徴(外観の変化)が出るまでの期間は樹齢や地域により異なりますが、成木では春から夏に感染したあと、9月〜10月ごろに枯れる
例が大半です。寒冷地では翌年の春に枯れることもあります。ここでは、長年の接種実験によって充分に確認された現象を中心に説明します。樹齢10年生前後
の木では接種から1〜2ヶ月で枯死することが確認されています。20年生以上ではややゆっくり進行したり、枝枯れで止まることもあります。小さな苗木への
接種では、組織の反応が速く、枯れ方が大木と異なることがあります。
感染後2週間程度、少数の線虫が
樹体内を移動
している時期に、幹では木部の水分通導機能(根から吸い上げた水を枝先まで上昇させる能力)を低下させる現象が起こります。この時期に伐倒して樹幹の断面
を見ると、写真-2Aのように白い斑点が見えます(Kuroda et al. 1988;
黒田1990ab)。肉眼で白く見えるのは、通導組織(仮道管、写真-1参照)に気体が充満したためです。酸性フクシンなどの色素液を樹幹の根元から吸い
上げさせると、水があがっている部分は染まりますが、白い部分はそのまま残ることから(写真-2B)、水分通導が一部で停止していることがわかりました。
気体による水分排除はキャビテーションあるいはエンボリズムと呼ばれます。通導組織が空になるという現象は健全な植物でも蒸散にともなって一時的に起こり
ますが、健全な場合はまた夜になると水で満たされるので萎れることはありません(Sperry & Tyree、 1988)。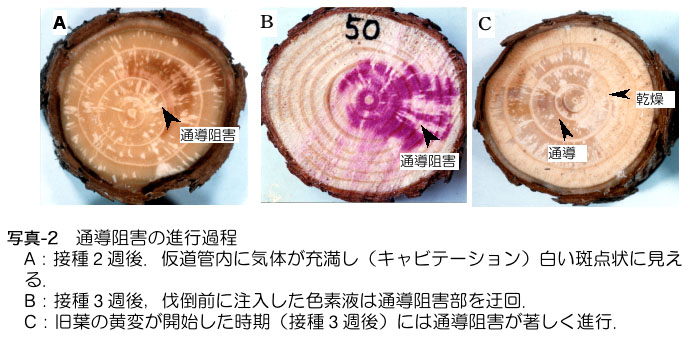
さて、材線 虫病の病徴を示して枯れたマツの材には青変菌など糸状菌類(カビ)が繁殖しているため、糸状菌が萎凋の真の原因ではないかという説もありました。しかし線 虫感染後の樹幹内の微生物の種類を調べると、葉が変色するなどの病徴が出る時期までは、病原性のない少数の微生物が健全木と同じ程度に検出されただけでし た。その後、樹液の流動が止まり、ほとんど枯れた段階で、キクイムシ類の侵入にともなって青変菌(Ceratocystis sp.)の感染がありました。青変菌の中には樹木を萎凋させる病原菌もありますが、線虫感染木では、枯れ始めたあとの感染であり、枯れる原因ではないこと が証明されました。(黒田・伊藤 1992)。
3. 萎凋現象を理解するために :マツ組織の構造と線虫の移動経路
樹幹横断面で気 体の充填した白い斑点が見える前、すなわち感染後の1〜2週間に、その原因となる生理的な変化が樹幹内で起こっていることになります。この変化について理 解するために、線虫の移動経路やマツの組織構造について、もう少し詳しく説明します。
自然感染の場合も接種の場合も、線虫の侵入場所と移動経路は同じです。線虫は枝の傷(マツノマダラカミキリの後食による噛み傷、ナイフによる切り傷)の上 に落ちると、皮層や木部にある樹脂道に、その切断部からもぐり込みます(図-1)。若枝の皮層内には縦(垂直)方向の樹脂道が、枝や主幹部の木部には垂直 方向と水平方向の樹脂道が存在します。3年生程度以上の枝や主幹部では皮層がなくなるため、線虫は皮層内の移動ができなくなり、移動経路には木部の樹脂 道が利用されるようになります(図-1、矢印)(市原、2000)。
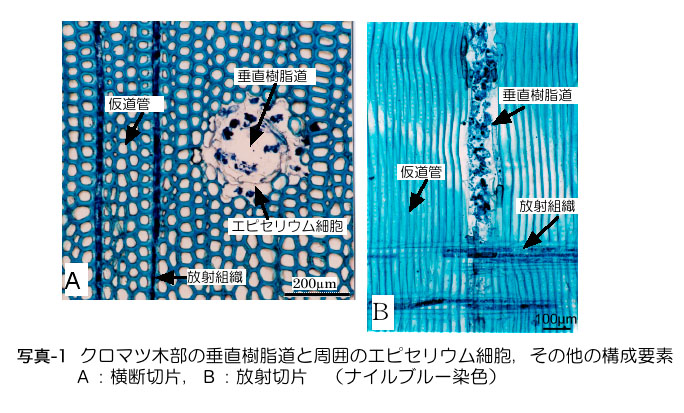
樹脂道とは細胞の間にできた筒状の隙 間(細胞間道)で、周 囲には樹脂を生産するエピセリウム細胞が取り巻いています(写真-1)。樹脂道内腔は直径が50〜60μmほどで、太さ10μm の線虫が中を通るだけのスペースは充分にあります。木部の垂直樹脂道の長さは数十cm程度あると推測されています。水平樹脂道と接して互いにつながってい るの で、移動の障害になるような構造はありません。針葉樹木部の大半は「仮道管」という水分通導経路となるソーセージ状の筒(長さ約1mm、太さ約30μm) が占めていますが(写真-1)、仮道管相互にあいている穴(壁孔:径1〜2μm)は線虫には小さすぎて移動経路に使えません。
樹脂道内には樹脂(松ヤニ)が溜まっているとは限らず、通常はほとんど空らしく、組織が傷ついた場合にエピセリウム細胞が迅速に樹脂生産を行います。枝の 傷口に落ちた線虫のほとんどは樹脂の中に閉じこめられてしまい、組織内に侵入することができません。接種実験では5000〜10000頭/本という多数の 線虫を接種しますが、実際に侵入できる線虫は1割程度です。
線虫は枝先から枝元へ、さらに主幹部へと樹脂 道の中を非常に速いスピードで移動します。接種後にクロマツを解体して線虫を 計数し、一日に150cm (約63mm/時間)も移動できることがわかりました(黒田・伊藤、1992)。線虫は重力の方向に移動しやすいようですが枝の先端方向にも移動できま す。また、全ての線虫が根にたどりつくのではなく、根の線虫が先に増えるとはかぎりません。移動の途中で各所にとどまる線虫もあります。移動経路になる 樹脂道が水平方向にも垂直方向にも多数分布すること、移動速度が速いことから、感染木では線虫分布は迅速に進みます。10年生程度のクロマツでは、 約1週間で全身分布することがわかっています(黒田・伊藤、1992)。直径が30cmもある大木ではもう少し時間がかかり、樹幹の一部に局在することも あります。当年生の苗では感染後すぐに全身に線虫が分布し、速く枯れます(Ichihara、 2001)。
この線虫 は雌雄があ り、樹幹内で交尾・産卵し増殖します。材線虫病に対する感受性が最も高い(一番枯れやすい)クロマツでも、感染から約3週間ほどは樹幹内の線虫数は少な く、 木部の乾燥重量1gあたり10頭程度までです(Kuroda et al. 1988)。線虫密度が100〜1000頭/gに上昇するのは、葉の変色が始まるころですから、そのころ幹の中が線虫の増殖に適した環境になると いう見方ができます。
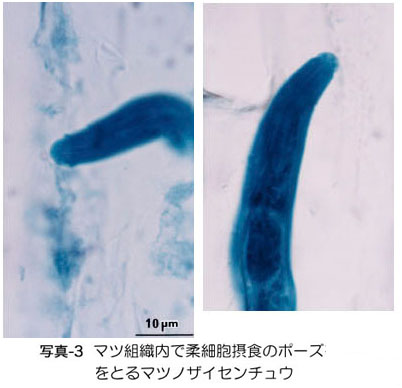
樹脂道の中で、線虫は柔細胞類(核も原形質も含む、生きている細胞)に口針(ストロー状の口)を挿入し (写真-3)、 内容物を酵素分解して吸収します(黒田・真宮1986)。摂食された柔細胞は壊死しますが、壊死する細胞は幹の組織の中のごく僅かであり、発芽直後の小さ な苗を除いて、「食べられたから枯れる」という状況には決してなりません。
「気体による急激な通導停止」 (写真-2) という現象について は、研究が進む前は予測がつかなかったため、昔は「線虫や松ヤニが仮道管(あるいは樹脂道)に詰まるために樹液が止まって枯れる」と説明されていました が、この説明は明らかに誤りです。移動中の線虫はまれに仮道管の中に頭を突っ込み、移動できずに止まることはありますが、通導阻害につ ながるほど頻繁ではありません。漏れた樹脂が仮道管に詰まるという現象は枯れかけた後の段階で見られますが、感染初期の通導阻害を説明するには無理があり ま す。
線虫が毒素を出すとか、感染したマツ組織が毒素を生産するという説もありました。しかし、樹木の細胞を多量に壊死させ枯死 に至らせるよ うな毒性物質は発見されていません。単に「未知の物質」程度の意味合いで「毒素」という言葉が用いられて、誤解をまねいています。「毒素」とい うことばには要注意です。
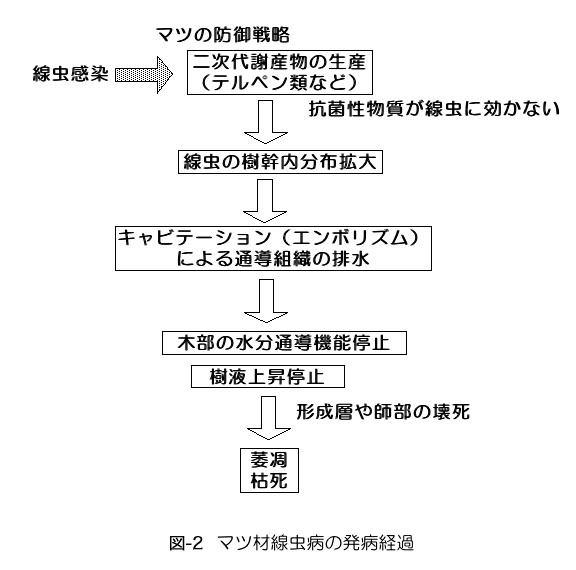
4. 通導阻害発生のメカニズム(図-2)
専門的な話になりますので、別項目で説明します。
5.マツ枯れに関わる誤っ た認識
マツの枯損は線虫によるのではなく、大気汚染が主原因であるという主張があります。接種 実験で「5千〜1万頭もの多量の線虫を接種して枯らしても原因とは言えない」と批判されることがあります。しかし、雌雄5対という少数の線虫接種で枯れる ことが確認されており(峰尾、1988)、接種頭数が多いから枯れたわけではありません。枯死のメカニズムを追求する研究では確実に接種木を枯らす必要が あるため、接種の1割しかマツ組織に侵入できない(前述)ことを前提に、接種頭数を多めに設定します。自然感染で、マツノマダラカミキリから1本のマツに 侵入する虫数にはかなりばらつきがあります。接種によって引き起こされた病気が自然感染による枯れと同じ経過をたどることを確認しており、異常な発病枯死 ではありません。
弱ったマツだから枯れやすいのではないか、健康なマツなら線虫が感染しても簡単に枯れないのではないかという 問いもあります。マツの生理的な活力の高さと感染後の抵抗性については、関連は充分に考えられます。特に「水分ストレス」といわれる水分の供給不足が病気 の進行に関わることはわかっており、トンネルや切り通しの崖ができると周囲でマツ枯れが起こりやすいという事例があります。しかし、酸性雨や大気汚染のひ どい地域でマツ枯損の発生が多いというデータ(相関関係があると表現される)だけから、「大気汚染がマツ枯れを促進する」という結論を導くことはできませ ん。大気汚染地域であるが、同時に著しい乾燥地である(例:瀬戸内地方の高速道路沿い)とか、マツ枯れの多発地で、線虫媒介者であるマツノマダラカミキリ の生息数が非常に多くなっている例など、他の要因を見落とすことになります。「健康なマツであれば線虫が感染しても大丈夫」ということはありません。アカ マツ、クロマツなど日本土着のマツは、材線虫病に対する感受性がきわめて高い種類で、健康なマツへの接種実験では7〜9割が枯れます。感染すれば枯れる確 率がきわめて高いということをご理解いただきたいと思います。
時には、「枯れ木から線虫が検出されない、 だから材線虫病で枯れたのではない」という意見があります。しかし、枯れた直後ではなく枯死後1年以上たってからの調査では、他の種類の線虫やいろいろな微生物 の繁殖が進み、マツノザイセンチュウが検出されないことがよくあります。また、幹の一カ所のみの調査では線虫が局在した場合に見逃すことがありますし、細 い枯れ枝では乾燥して線虫が居なくなっていますから、線虫は検出できません。材線虫病の感染木では枯れの進み方が特徴的で、関東以西では感染木の多くは9 月頃に急激に葉が赤変すること、東北などの冷涼な地域では、翌年1月から4月ごろに萎凋症状を示す例も多いことを知っているなら、他の原因による枯損との 違いは見当がつきます。さらに組織内の線虫の種類を専門家が確認すれば診断できます。日本では、北海道以外で見られるマツ枯れは、その大半がマツ材線虫病 によるものです。
6.抵抗性とは →次のページ
引用文献
- Fukuda, K. (1997). Physiological process of the symptom development and resistance mechanism in pine wilt disease. J. For. Res. 2: 171-181.
- 二井一禎・古野東州(1979).マツノザイセンチュウに対するマツ属 の抵抗性.京大演習林報告 51,23-36.
- 市原 優(2000).マツノザイセン チュウの通り道.森林総合研究所東北支所たより 462:1-4. Ichihara, Y.,
- Fukuda, K., and Suzuki, K. 2000. Early symptom development and histological changes associated with migration of Bursaphelenchus xylophilus in seedling tissues of Pinus thunbergii. Plant Disease 84, 675-680.
- 黒田宏之・黒田慶子・高井一也・ 鈴木敏雄 (1999).マツ枯損防止のための新戦略構築.H9-10年度科学研究補助金研究成果報告書(研究課題番号09556037)84pp.& amp; amp; amp; amp; amp; nbsp;
- Kuroda, K. (1989).Terpenoids causing tracheid-cavitation in Pinus thunbergii infected by the pine wood nematode (Bursaphelenchus xylophilus). 日本植物病理学会報 55,170-178.
- 黒田慶子 (1990).マツ材線虫病の発病および病徴進展にかかわる通水阻害.日本農芸化学会誌64:1258-1261.
- 黒 田慶子 (1990).特集 :線虫学(5) マツノザイセンチュウ感染によるマツの枯損機構.植物防疫 44(12), P539-542. Kuroda, K. .Mechanism of cavitation development in the pine wilt disease. Eur. J. For. Path. 21, 82-89.(1991)
- 黒田慶子 (1995).マツ材線虫病の発病機構と抵抗性機構 -抵抗性樹種における発病阻止要因の検討-.森林総合研究所所報 85, 8-9.
- 黒田慶子 (1996).樹病:病原微生物の感染戦略と樹木の反応.木材保存 22(4),205-214. 黒田慶子 (1998).樹木の通水機能と萎凋病.森林保護 267,36-38.
- 黒田 慶子(1999).樹木医学(鈴木和夫編)2.2樹木の構造と機能.朝倉書店,57-82.
- 黒田慶子・伊藤進一郎 (1992).クロマツに侵入後のマツノザイセンチュウの動きとその他の微生物相の変遷.日本林学会誌, 74(5), 383-389.
- 黒田慶子・真宮靖治 (1986) .マツノザイセンチュウの無菌クロマツ稚苗内における行動.日本林学会大会論文集 97回 P471-472.
- Kuroda, K., Yamada, T. and Ito, S. (1991). Bursaphelenchus xylophilus induced pine wilt: Factors associated with resistance. Eur. J. For. Path. 21, 430-438.
- 黒田慶子・山田利博・伊藤進一郎 (1991).アカマツにおけるマツ材線虫病の進行と通水異常.日本林学会誌 73(1):69-72.
- Kuroda, K., Yamada, T., Mineo, K. and Tamura, H (1988).Effects of cavitation on the development of pine wilt disease caused by Bursaphelenchus xylophilus. 日本植物病理学会報,54,606-615.
- 峰尾一彦(1988).マツノザイセンチュ ウの週数接種とマツ苗木の発病.森林防疫 37:35-36.
- 島地 謙・須藤彰司・原田 浩(1976)木材の組織.森北出版,291pp.
- Sperry, J.S., Tyree, M.T. (1988). Mechanism of water stress-induced xylem embolism. Plant Physiol. 88: 581-587.
- 戸 田忠雄・寺田貴美雄(2001).林木育種のプロジェクト(3) ---マツノザイセンチュウ抵抗性育種事業---.林木の育種 198:39-43.