マツ樹幹内で起きていること
---マツ材線虫病の発病機構と抵抗性に関する研究より- ---
黒田慶子
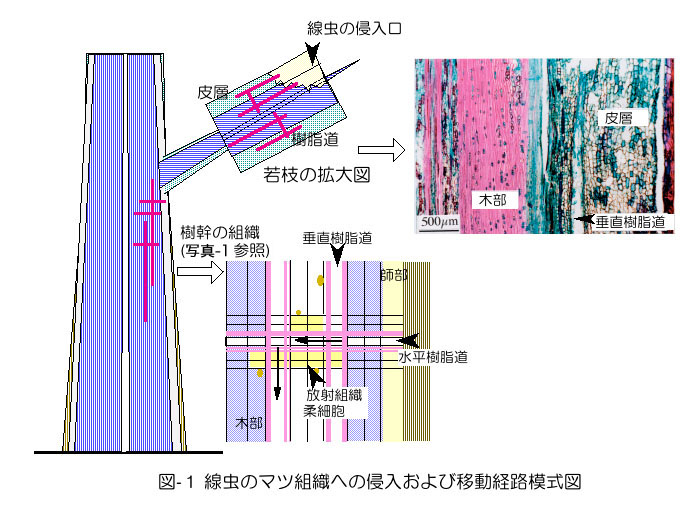
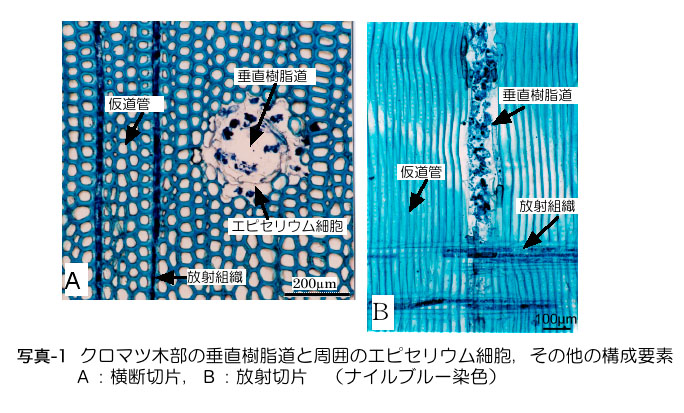
マツ材線虫病の発病および萎凋のメカニズムは,かなり細部まで説明できるようになった.しかし依然として「発病機構はまだわ かっていない」と一言で片づけられることが多く,「マツノザイセンチュウ(以下線虫)は病原体でないかも知れない」と疑いを持たれる原因ともなっている. 発病機構については何度か解説の機会を得ているが(黒田,1990ab,1996,1998),ここでは最新の情報を紹介するのではなく,造園や林業,行 政などの現場の疑問を念頭に基本的な事柄を整理して解説する.発病機構とは,「線虫の感染からマツが萎凋するまで」の道筋の説明である.樹幹の外から見え ない現象がイメージできるように,マツの組織の立体構造や機能について,図や写真により解説を加えた(図-1,写真-1).
線虫が感染したクロマツやアカマツに最初に見られる病徴(外観の変化)は
旧葉の変色と,時には頂端部の葉の萎れ(下垂)である
が,樹幹の中ではそれ以前に大きな変化が起こっている.感染から病徴が出るまでの期間は樹齢により異なるが,成木で1〜2か月という認識が一般的であろ
う.ここでは,接種実験によって十分に確認のできた樹齢10年生前後の例を中心に樹幹内の現象を説明する.20年生以上ではややゆっくり進行したり,局部
的に現象が進むこともある.数年生以下の苗木への接種では,組織の反応がかなり異なることも認識する必要がある.
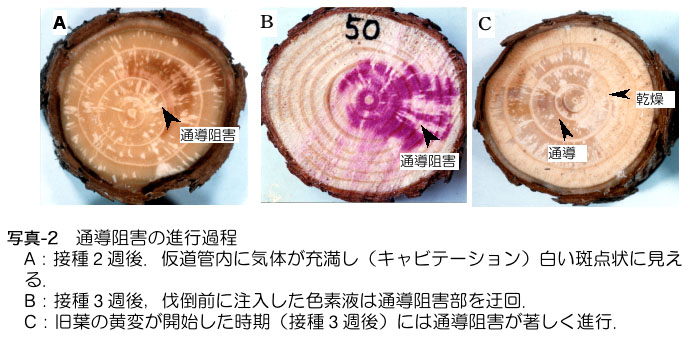
感染後2週間程度,少数の線虫が樹体内を移動している時期に,主幹部では木部の水分通導機能(根から吸い上げた水を枝先まで上昇させる能力)を低下させ る現象が起こっている.この時期に伐倒して樹幹の断面を見ると,写真-2Aのように白い斑点が見える(Kuroda et al., 1988; 黒田1990ab).肉眼で白く見えるのは,通導組織(仮道管,写真-1参照)に気体が充満しているためである.酸性フクシンなどの色素液を樹幹基部から 吸い上げさせると,白い部分はそのまま残ることから(写真-2B),水分通導が一部で停止していることがわかる.気体による水分排除はキャビテーションあ るいはエンボリズムと呼ばれる.通導組織が空になるという現象は健全な植物でも蒸散にともなって一時的に起こるが,健全な場合はまた水で満たされるので萎 れにはつながらない(Sperry & Tyree, 1988).しかし線虫感染木では,このように気体が充填した部位が再度水で満たされることはなく,白い部分が増えて,水分通導の能力が急速に低下する. 通導阻害部位の拡大(写真-2C)と,旧葉の変色に始まる病徴発現とは時期がほぼ一致しており,著しい通導低下が萎凋につながっていることは明らかにであ る.10年生前後のクロマツでは線虫接種後約4〜5週間,アカマツでは6週間程度で主幹部の水分通導がほぼ全面的に停止する.さらに木部が乾燥するととも に形成層や師部の壊死が進行し,樹木全体が枯損の様子を示す(Kuroda et al., 1988; 黒田ら1991).このような急激な木部の乾燥は,切り枝を大気中に放置しても見られない現象である.
さ て,材線虫病の病徴を示して枯れたマツの材には青変菌など糸状菌類(カビ)が繁殖しているため,糸状菌が萎凋の真の原因ではないかという説もあった.しか し線虫感染後の樹幹内の微生物相の変遷を見ると,病徴が出る時期までは,少数の微生物が健全木と同じ頻度で検出されるのみで,これらは病原性を持たない. 通導阻害が顕著になり枯死に進む過程で,キクイムシ類の集中的樹幹穿入にともなって青変菌(Ceratocystis sp.)が感染していた.青変菌の中には樹木を萎凋させる病原菌も存在するが,線虫感染木については発病への関与が否定された.(黒田・伊藤, 1992).
3. 萎凋現象を理解するために :マツ組織の構造と線虫の移動経路
樹幹横断面で気体の充填した白い斑点が見える前,すなわち感染後の1〜2週間に,その原因となる生理的な変化が線虫の活動に よって樹幹内で起こっていることになる.その現象について説明するまえに,マツ樹幹内での線虫の移動経路やマツの組織構造についてふれておきたい.
& nbsp; 自然感染の場合も接種の場合も,線虫の侵入経路は同じである.線虫は枝の傷(マツノマダラカミキリの後食による噛み傷,ナイフによる切り傷) の上に落ちると,皮層や木部にある樹脂道に,その切断部からもぐり込む(図-1)注1).若枝の皮層内には縦(垂直)方向の樹脂道が,枝や主幹部の木部に は垂直方向と水平方向の樹脂道が存在する.3年生枝程度以上の枝や主幹部では周皮形成や皮層の脱落によって,線虫は皮層内の移動ができなくなり,線虫の移 動経路には木部の樹脂道が利用されるようになる(図-1,矢印)(市原,2000).
樹脂道とは細胞の間 にできた筒状の隙間(細胞間道)で,周囲には樹脂を生産するエピセリウム細胞が取り巻いている(写真-1).樹脂道内腔は直径数十μmあり,太さ10μm の線虫が中を通るだけのスペースは充分にある.木部の垂直樹脂道の長さは数十cm程度あると推測されており,水平樹脂道と接して互いにつながっているの で,移動の障害になるような構造はない.針葉樹木部の大半は「仮道管」という水分通導経路となるソーセージ状の筒(長さ約1mm,太さ約30μm)が占め ているが(写真-1),仮道管相互にあいている穴(壁孔:径1〜2μm)は線虫には小さすぎて移動経路に使えない.
樹脂道内には樹脂(松ヤニ)が溜まっているとは限らず,通常はほとんど空らしく,組織が傷ついた場合にエピセリウム細胞が迅速に樹脂生産を行う.枝の傷 口に落ちた線虫の多くは樹脂の中に閉じこめられ,組織内に侵入するのは困難である.接種実験では5000〜10000頭/本という多数の線虫を接種する が,侵入できる線虫は1割程度である(未発表).接種後の線虫増殖について検討する場合は,感染時の線虫数イコール接種頭数ではないことに注意する必要が ある.
線虫は枝先から枝元へ,さらに主幹部へと樹脂道の中を非常に速いスピードで移動する注2).接種後 にクロマツを解体して線虫を計数し,一日に150cm (約63mm/時間)も移動できることがわかった(黒田・伊藤,1992).線虫は重力の方向に移動しやすいようであるがシュートの先端方向にも移動す る.また,全ての線虫が根にたどりつくのではなく,根の線虫密度が先に高くなるとは限らない(未発表).移動の途中で各所にとどまる線虫もいる.水平方向 にも垂直方向にも移動経路の樹脂道が多数分布すること,移動速度が速いことから,感染個体の木部では,線虫分布(分散)は迅速に進み,10年生程度のクロ マツでは,約1週間で確実に全身分布する(黒田・伊藤,1992).直径が30cmもある大木ではもう少し時間がかかり,樹幹の一部に局在することもあ る.当年生の苗では感染当初から線虫密度が高く,病徴進展が非常に速い(Ichihara, 2001).感染から病徴発現に至るまでの時間(日数)は,線虫の分布に要する時間に依存する部分が大きいと思われる.
樹幹内 の線虫は交尾・産卵し増殖する.材線虫病に対する感受性が最も高いクロマツで,感染から約3週間は樹幹内の線虫密度は低く,木部の乾燥 重量1gあたり10頭程度〜100頭未満である(Kuroda et al. 1988).線虫密度が1000頭/g以上の桁に上昇するのは,萎凋症状が出始めるころで,すなわち線虫の増殖に適した環境になった時期であるという解釈 もできる
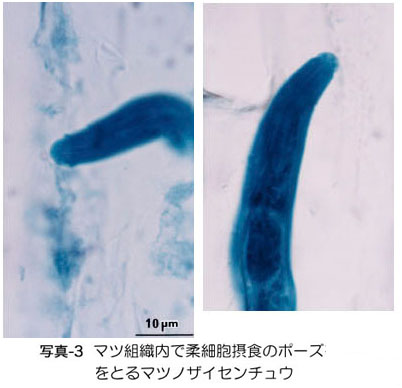 .
.& nbsp; 樹脂道の中で,線虫は柔細胞類(核も原形質も含む,生きている細胞)に口針(ストロー状の口) を挿入し(写真-3),内容物を酵素分解して吸収する(黒田・真宮,1986).摂食された柔細胞は膨れたり壊死したりするが,摂食によって壊死する細胞 は樹体のごく僅かであり,発芽直後の小さな苗を除いて,枯れる原因には決してならない.
「気体による通導 阻害」という現象については,研究が進む前は予測がつかなかったため,枯損理由は「線虫や松ヤニで仮道管(あるいは樹脂道)が詰まるた めに樹液上昇が止まって枯れる」と説明されていた時期があるが,この説明は明らかに誤りである.移動中の線虫は樹脂道から仮道管の中に頭を突っ込み,それ 以上移動できずに止まることはあるが,そのような事例の発生頻度は通導阻害につながるほどではない.また,漏れ出た樹脂が仮道管に詰まって水の流れを悪く するという考え方も,感染初期の通導阻害を説明するには無理がある.
なお,線虫が毒素を出す,感染したマツ組織が毒素を生産す るという説があるが,樹木の細胞を多量に壊死させ枯死に至らせる毒性物質は発見されていない.こ のような「未知の物質」程度の意味合いで「毒素」という言葉を用いると,誤解をまねくことになる.時には,セルラーゼなど分解酵素や樹木組織で生産された 二次代謝成分の一部を広義の毒素と解釈する例があるが,安易に「毒素」と表現するのは避けるべきである。
& nbsp;注1 : 2〜3年生程度までの若枝には外樹皮はなく,表皮,皮層があり,その内側に二次師部(内樹皮)が形成されている(黒田,1999).枝が太くなるにつれて 皮層は脱落し,外樹皮と内樹皮という構造になる(図-1参照).
注2 :内樹皮(師部)には垂直樹脂道は存在しない.皮層の樹脂道を移動している線虫は枝元の太い部分や主幹部(皮層が脱落して樹皮が形成された部位)に至る前 に経路を変え,水平樹脂道を通って木部の樹脂道に移動する必要がある.皮層内の移動は袋小路で止まる割合が高いと思われる.
4. 通導阻害発生のメカニズム
では,感染木では何が原因で回復不能のキャビテーションが起こり,通導停止へと進行するのだろうか.まだ推測の部分もあるが, 部分的には説明が可能である.明らかなことは,感染個体では樹液の流動が非常に途切れやすくなっていることである(黒田1990).木部樹液の性質が変化 したと考えるのが今のところは妥当である.あるいは,柔細胞の能力が弱まって樹液上昇を助ける機能が働かなくなった可能性もある.
健全な針葉樹が根から水を吸収して葉先まで供給できるのは,葉から水分が蒸発するにつれて仮道管内の樹液が引っ張り上げられるからであるが, この引っ張りに対して樹液が耐えられなくなると,経路の途中(仮道管)で気体が発生し,仮道管内が気体に置き換わってしまう(Sperry & Tyree, 1988; Kuroda, 1991).これは空気が外界から取り込まれるのではなく,沸騰に似た現象であるため理解されにくいが,実験的に意外に簡単に起こせる現象である注3). 本来,木部樹液は純粋な水に近いと考えられており,この中に混ざった物質により液の表面張力が低下した場合に気体発生(キャビテーション)が促進される (Sperry and Tyree1988).感染にともなって生成した物質が樹液内に混じった場合,同様の現象が起こりうる.マツノマダラカミキリの誘因物質として知られるモ ノテルペンは,線虫接種の数日後からマツ組織内で増加するが,これは表面張力を低下させる物質である (Kuroda, 1989; 1991).他にも表面張力低下に関わる物質が生成したり,気体の発生が促進されることもあり得る.健全木ではキャビテーションが起こっても再度水が流入 して通導能力は回復するが,線虫感染木では水の再流入はなく,樹液上昇は回復しない.感染後1〜2週間後から,樹脂道の周囲などに油滴(揮発性テルペンを 含む)の漏出が顕微鏡で確認できることから(Kuroda, 1989),疎水性の物質が壁孔膜や細胞壁に付着することも,通導回復を妨げる理由の一つではないかと推測している.原因の特定にはさらに追求が必要であ る.
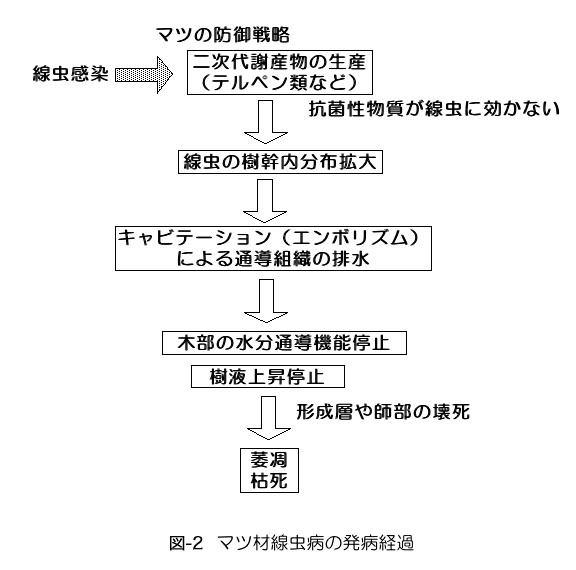
& nbsp; 「木部樹液があがらなくなる現象」がどのように起こっているかについては,以上のように経過を説明す ることが可能になった.樹液の供給が減少するにつれて,形成層など生きている組織の乾燥や壊死が進み,それに引き続いて,葉の萎れや変色などの病徴(外見 的な症状)が進展する.線虫が感染するとなぜ枯れるかという問いには,図-2に示すように,「線虫の移動や摂食活動による刺激→マツ細胞の代謝異常(二次 代謝産物の生成注4)→木部樹液内で気泡発生,通導組織から排水→通導阻害→水分供給停止→枯死」という回答で,ひとまず十分と考えている. 注3 :針のない注射器に水を半分入れ,先を塞いでピストンを引くと,簡単に気体が発生する.空気が入ったわけではないので,手を離すと気体は消失する. http://cse.ffpri.affrc.go.jp/keiko/hp/embolism.htmlに,顕微鏡下でマツ(切片)の仮道管にキャビ テーションが起こる様子をビデオ画像で紹介している. 注4:二次代謝産物は樹木の心材化の際,あるいは外敵の侵入に対する抵抗反応として樹体内で生産されるテルペン類やフェノール性の物質などの総称である. 抗菌性を示す物質も含むが,本病に感受性の高いマツの場合,線虫の活動を阻害する効果はみられないようである.
5.マツ枯れに関わる誤っ た認識
マツの枯損は線虫によるのではなく,大気汚染が主原因であるとの主張がまだ続いている.接 種実験については 「10000頭もの多量の線虫を接種しなければ枯れないのでは,病原力が強いとはいえない」と批判される.しかし,雄雌5対という少数の線虫接種で枯損す ることが確認されており(峰尾,1988),接種頭数が多いことを理由に線虫の病原性を疑うことはできない.枯死メカニズムを追求する研究では確実に同時 に枯らすことが不可欠であるため,接種頭数の1割しかマツ組織に侵入できない(前述)ことを念頭に,接種頭数を5000〜10000頭と多めに設定する. 自然感染において,マツノマダラカミキリからマツ一個体に侵入できる線虫の頭数は明らかではないが,線虫接種による枯損経過は自然感染による枯損経過と同 じであることを確認しており,接種実験で異常な発病枯死を起こしてはいない.
弱ったマツだから枯れやすい のではないか,健康なマツなら線虫が感染しても簡単に枯れないのではないかという問いも多い.マツの生理的な活力の高さと感染後の抵抗性については,関連 は充分に考えられる.特に水分ストレス(水分の供給不足)の影響は推測されており,トンネルや切り通しの崖ができるとマツ枯損が目立つなどの事例がある. しかし,酸性雨や大気汚染とマツ枯損発生地との相関関係から結論を急いではいけない.大気汚染地域であってしかも著しい乾燥地である(例:瀬戸内地方の高 速道路沿い)とか,恒常的にマツ枯損が続いていて,線虫媒介者であるマツノマダラカミキリの生息数が多い地域など,他の要因を見落とすことになる.「健全 であれば線虫が感染しても大丈夫」ということはない.アカマツ,クロマツなど日本土着のマツは,本病に対する感受性がきわめて高い種であり,感染すれば枯 れる確率が高いことを念頭に置かなければならない.
枯れたマツからマツノザイセンチュウが検出されない, マツ枯れは線虫によらない枯損が多いのではないかという意見がある.枯れた直後に調査しなかった場合,枯死後1年程度で他の種類の線虫に交代してしまうた め,検出されないことがよくある.また,線虫を分離する試料として細い枯損枝のみを用いた場合,乾燥のため線虫は存在しないことが多く,幹の一カ所のみの 調査では線虫が局在した場合に見逃すことがある.日本では,北海道以外で見られるマツ枯れは,すべてではないがその大半は材線虫病による枯損である.材線 虫病による枯損は病徴の出方(枯れ方)が特徴的で,関東以西では感染木の多くは9月頃に急激に葉が赤変すること,冷涼な地域では,翌年1月から4月ごろに 萎凋症状を示す例も多いことを知っているなら,他の原因による枯損との差は認識できる.組織内の線虫の種類を確認すれば診断は難しくない.
& nbsp; この線虫は樹木細胞を食べることにより枯らしているのではないこと,マツの組織自身が「木部樹液の流れを止める」現象を起こしていることが認 識されれ ば,線虫の病原性に関する疑いは晴れるはずである.
6. 抵抗性とは
マツ材線虫病で取り組むべき課題がかなり残っているのは,抵抗性に関する部分である.マツノザイセンチュウは外国産の線虫であ り,日本産のマツはこの線 虫とのつきあいが短いため感受性が高いと言われる(二井,古野 1979).北米産のテーダマツは抵抗性が非常に高く,接種により枯れる率が低い (100%枯れないわけではない).「なぜ枯れないか」を明らかにすれば,防除に役立つ情報が得られるだろうという期待がある.抵抗性の種として知られる マツの組織内では線虫の移動と増殖が活発ではないことがわかっている(Kuroda et al. 1991,黒田1995).
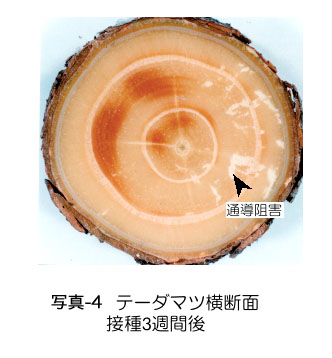
抵抗性のマツであっても,線虫が樹幹組織内に侵入して移動することは可能である.接種2〜3週間後には,テーダマツ樹幹にも感受性の高いクロ マツなどと同様に白い斑点(通導阻害部位)が認められる(写真-4).しかしその範囲は非常に狭く,樹体全体が水分供給不足となるほどには進展しない (Kuroda et al. 1991).興味深いのは,このように抵抗性の種でも線虫の影響があることである.通導阻害の発生が線虫の移動摂食した狭い範囲に限定されていたため病徴 発現には至らなかったのであり,「感染したが病徴は進展しなかった」と言える.テーダマツでは接種枝のみ枯死する場合があり,多数の線虫が存在すれば,病 徴進展につながることを示している.テーダマツ組織には線虫の活動を阻害する物質が存在,あるいは生成するのではないかと推測されるが,候補として挙げら れたβミルセンの関与については肯定的なデータが得られていない(Kuroda et al., 1991).
感受性種である アカマツ,クロマツについては,抵抗性個体の選抜と育種が林木育種センターや多くの府県で進められており(戸田・寺田,2001),抵抗 性発現のメカニズムについても追求し始めた.抵抗性発現に関わる物質の特定は困難であろうが,線虫の移動を遅らせるような現象が認められるなら,選抜家系 について抵抗性の程度の評価をすることも可能であろう.一方では,線虫の活動阻害効果のある物質をマツ樹体内に誘導するなどの方法で,抵抗性を付与すると いう考え方もある(黒田ら1999).
7. おわりに
以上の発病機構に関する解説は,これまでわかった中で最も単純な部分を簡潔に説明したものである.原著論文を網羅して引用できないので,解説 文を中心に引用した.
文献の一部はhttp: //cse.ffpri.affrc.go.jp/keiko/hp/kuroda.htmlに掲載してい る.樹木の構造と機能については,島地ら(1976)や黒田(1999)も参照していただきたい.
本稿で 紹介した現象やそれに付随して起こる事柄,線虫の挙動などについて,綿密な研究が進んでおり,詳細なデータが蓄積されつつある(Fukuda, 1997).本稿に引き続き解説され,材線虫病に関する理解がさらに深まることを望んでいる.
引用文献
- Fukuda, K. (1997). Physiological process of the symptom development and resistance mechanism in pine wilt disease. J. For. Res. 2: 171-181.
- 二井一禎・古野東州(1979).マツノザイセンチュウに対するマツ属 の抵抗性.京大演習林報告 51,23-36.
- 市原 優(2000).マツノザイセン チュウの通り道.森林総合研究所東北支所たより 462:1-4. Ichihara, Y.,
- Fukuda, K., and Suzuki, K. 2000. Early symptom development and histological changes associated with migration of Bursaphelenchus xylophilus in seedling tissues of Pinus thunbergii. Plant Disease 84, 675-680.
- 黒田宏之・黒田慶子・高井一也・ 鈴木敏雄 (1999).マツ枯損防止のための新戦略構築.H9-10年度科学研究補助金研究成果報告書(研究課題番号09556037)84pp.& nbsp;
- Kuroda, K. (1989).Terpenoids causing tracheid-cavitation in Pinus thunbergii infected by the pine wood nematode (Bursaphelenchus xylophilus). 日本植物病理学会報 55,170-178.
- 黒田慶子 (1990).マツ材線虫病の発病および病徴進展にかかわる通水阻害.日本農芸化学会誌64:1258-1261.
- 黒 田慶子 (1990).特集 :線虫学(5) マツノザイセンチュウ感染によるマツの枯損機構.植物防疫 44(12), P539-542. Kuroda, K. .Mechanism of cavitation development in the pine wilt disease. Eur. J. For. Path. 21, 82-89.(1991)
- 黒田慶子 (1995).マツ材線虫病の発病機構と抵抗性機構 -抵抗性樹種における発病阻止要因の検討-.森林総合研究所所報 85, 8-9.
- 黒田慶子 (1996).樹病:病原微生物の感染戦略と樹木の反応.木材保存 22(4),205-214. 黒田慶子 (1998).樹木の通水機能と萎凋病.森林保護 267,36-38.
- 黒田 慶子(1999).樹木医学(鈴木和夫編)2.2樹木の構造と機能.朝倉書店,57-82.
- 黒田慶子・伊藤進一郎 (1992).クロマツに侵入後のマツノザイセンチュウの動きとその他の微生物相の変遷.日本林学会誌, 74(5), 383-389.
- 黒田慶子・真宮靖治 (1986) .マツノザイセンチュウの無菌クロマツ稚苗内における行動.日本林学会大会論文集 97回 P471-472.
- Kuroda, K., Yamada, T. and Ito, S. (1991). Bursaphelenchus xylophilus induced pine wilt: Factors associated with resistance. Eur. J. For. Path. 21, 430-438.
- 黒田慶子・山田利博・伊藤進一郎 (1991).アカマツにおけるマツ材線虫病の進行と通水異常.日本林学会誌 73(1):69-72.
- Kuroda, K., Yamada, T., Mineo, K. and Tamura, H (1988).Effects of cavitation on the development of pine wilt disease caused by Bursaphelenchus xylophilus. 日本植物病理学会報,54,606-615.
- 峰尾一彦(1988).マツノザイセンチュ ウの週数接種とマツ苗木の発病.森林防疫 37:35-36.
- 島地 謙・須藤彰司・原田 浩(1976)木材の組織.森北出版,291pp.
- Sperry, J.S., Tyree, M.T. (1988). Mechanism of water stress-induced xylem embolism. Plant Physiol. 88: 581-587.
- 戸 田忠雄・寺田貴美雄(2001).林木育種のプロジェクト(3) ---マツノザイセンチュウ抵抗性育種事業---.林木の育種 198:39-43.